


Све је резултат његошевског „согласија општег”, које се своди на то да су у природи могући само они системи који су детерминисани најбољом могућом – симетријом, пропорцијом и хармонијом, у чијој основи је златни пресек. Како је пре четрдесет гдина ђаво дошао по своје?

Проф. др Милоје М. Ракочевић
Ни у чему што нам се дешава
нисмо сами, ни први ни једини.
Иво Андрић (Знакови поред пута)
Пре четрдесет година дошао је ђаво по своје! Пробудио сам се једног јутра до дна свог бића погођен необичном загонетком природе. И како тада, тако и данас, ђаво ми не да мира – не престајем да се питам зашто је то са животом баш тако, сводиво на само два питања (оба питања са по једним потпитањем): 1. Сви гени свега живог, од бактерије до човека, саздани су од само четири мала молекула, четири специфичне азотне, то јест нуклеотидне базе, две пиримидинске (једнопрстене) и две пуринске (двопрстене); и потпитање: зашто баш та четири молекула? 2. Сви протеини свега живог саздани су од тачно двадесет аминокиселина – двадесет аминокиселинских молекула; и потпитање: зашто баш тих 20 молекула?
[Свака од две пиримидинске базе (тимин/урацил, цитозин) и две пуринске (аденин, гуанин) сједињава се са по једним молекулом шећера рибозе, или дезоксирибозе, и са по једним молекулом фосфорне киселине, па тако настаје нови, још увек релативно мали молекул – нуклеотид, основна градивна јединица макромолекула рибонуклеинске киселине (РНК, са урацилом), односно дезоксирибонуклеинске киселине (ДНК, са тимином)].[1]
Да би се могло одговорити на два основна питања – зашто 4 и зашто 20 – можда се и не мора знати хемија, али да би се могло одговорити на два потпитања – зашто баш та и таква 4 нуклеотидна молекула, и зашто баш тих и таквих 20 аминокиселина – мора се бити хемичар, што се мени, више случајним неголи намерним избором студија, и догодило (иако у гимназији хемију нисам ни волео).
На путу ка могућим одговорима на постављена питања постоји једно „али”. Постављена питања спадају у тзв. фундаментална питања, а код њих не функционише форма зашто, већ једино како. Не можемо одговорити на питање зашто је то тако, већ једино на питање – како стоје ствари у вези са постављеним питањем. Не можемо знати зашто је то тако да универзум постоји, а није тако – да не постоји. Не знамо зашто је то тако да су хемијски елементи саграђени од атома, а није тако да су саграђени на неки други (и који?) начин.
[1] Број атома у четири молекула који граде информациону рибонуклеинску киселину, iRNK, кореспондентно са Таблицом генетског кода (Табела 1), износи: U = 12, C = 13, A = 15, G = 16; односно, у паровима: UG = 28 x 1 и CA = 28 x 1. Кад поприме форму нуклеотида, тада је број атома следећи: U = 34, C = 35, A = 37, G = 38; односно, у паровима: UG = 36 x 2 и CA = 36 x 2 (упоредити: Rakočević, 1997а / 1997b, Solutions 25-29, pp. 62-63; www.rakocevcode.rs). Поставља се питање: да ли је ово куриозитет и коинциденција, или је у питању извесна природно-кодна суштина кад суме броја атома по паровима корспондирају са квадратом првог савршеног броја (6) и првим степеном другог савршеног броја (28)? [Савршен је онај број који је једнак суми својих чинилаца.]
Јединствен број
Са овим сазнањем, погледајмо како стоје ствари у вези са два основна питања. Прво питање тиче се броја 4. Тај број је специфичан и уникатан у том смислу што поседује једно својство које не поседује ниједан други број у низу природних бројева. Посматран као целина, тај број се са својом половином (симетрија!) налази у релацији једнакости кроз три операције: 2 + 2 = 2 x 2 = 22 = 4. Осим тога, релација једнакости остварује се и кроз операцију инверзије, и то у границама скупа {2, 4}, где је 24 = 42; а, такође и у границама проширеног скупа, проширеног следећим парним бројем, бројем шест {2, 4, 6}, где је 26 = 43 = 64; а, управо толико има кодовних „речи” (кодона) у генетском кôду. Све ово нас доводи у везу и са другим значајним релацијама, кроз одговарајућа математичка уопштавања.
Тако једноставна таблица, а у њој
садржана комплетна прича о једном
чудесном језику – језику гена;
прича о четворословној азбуци из
које настају 64 трословне речи.
А најзначајнија веза, непосредно видиљива кроз та уопштавања, јесте она која представља непосредну кореспонденцију израза за генетски, и израза за хемијски кôд, како је и приказано у прегледу 1 (у релацији са прегледом 2): mn = nm / n2 = 2n. Наравно, ова веза се може уочити само под условом да се периодни систем хемијских елемената (ПСЕ) представи на адекватан (истинит) начин, како сам и показао још пре 26 година (Ракочевић, 1991).[2]
[2] У питању је Tabela 4.1 у наведеном раду (Ракочевић, 1991), која се налази и у књизи (Rakočević, 1997b), као Табела 18, стр. 180 (www.rakocevcode.rs). Главно у тој Таблици ПСЕ јесте то што је 14 лантанида распоређено у 14 група, на исти начин како је то учинио Мендељејев (Кедров, 1977, Таблица 18, стр. 188). У том смислу, остаје нејасно зашто већ деценијама IUPAC (International Union of Pure and Applied Chemistry), у чијој компетенцији је, поред свега другог у хемији, и номенклатура ПСЕ, игнорише Мендељејева, (стављањем свих 14 лантанида, заједно са лантаном, у трећу групу), игноришући тиме и основни закон хемије (који проистиче из закона периодичности), према којем сваки непарни елеменат мора бити у непарној, а парни у парној групи. [IUPAC: „Project No.: 2015-039-2-200; Start Date: 18 December 2015; Division Name: Inorganic Chemistry Division; Division No.: 200. Objective: This project will deliver a recommendation in favor of the composition of group 3 of the periodic table as consisting either of: 1. the elements Sc, Y, Lu and Lr, or 2. the elements Sc, Y, La and Ac. The task group does not intend to recommend the use of a 32-column periodic table or an 18-column. This choice which is a matter of convention, rather than a scientific one, should be left to individual authors and educators.“ (Оба подвлачења – лутецијума и лантана – јесу моја, да би се истакло да IUPAC својим пројектом жели да разреши наводну дилему који од ова два елемента треба да буде уписан у трећу групу?!)]
Друго питање тиче се броја 20, а да би се сагледала његова специфичност и уникатност нужно је да се таблица множења у декадном бројевном систему представи на адекватан (истинит) начин, када је непосредно очигледно да се број 20 у форми 20-20 налази тачно на средини таблице (видети уоквирени текст).
Што се тиче могућег одговора на два потпитања – зашто баш та 4 нуклеотидна молекула, и баш тих 20 аминокиаелинских молекула – потребно је написати бар још једну, добро аргументовану, макар и малу студију, што овим чланком делимично и чинимо.
*
Те, сада давне, 1977. године, Таблица генетског кôда (табела 1) постојала је тек десетак година, мада је предисторија њеног настанка већ увелико настањивала све средњошколске уџбенике широм света. Тако једноставна таблица, а у њој садржана комплетна прича о једном чудесном језику – језику гена; прича о четворословној азбуци из које настају 64 трословне речи, за које би математичари рекли да су то варијације треће класе (са понављањем) из скупа од четири елемента.
И већ у овоме се види јединственост, тј. уникатност генетског кода. Једино у овом (могућем кодогеном) случају важе два основна мендељејевска принципа – принцип минимума промене, и принцип континуитета: азбука је четворословна, реч трословна, корен речи двослован, старт једнослован. А, ако би азбука била, на пример, петословна, поштовањем два наведена принципа речи би морале бити четворословне, али тада постоји неизвесност за корен речи – да ли је са три, или са два слова?
Бројеве по дијагонали таблице множења (Табела 2): 0, 2, 6, 12, 20, 30, … , препознајемо и као вредности за q у једначини генералисаног златног пресека x2 + px = q (q = 0, 2, 6, 12, 20, 30, …) у скупу фамилије „металних пресека” (Spinadel, 1998, 1999; Rakočević, 2004b), где се решења тих једначина генеришу из скупа природних бројева: (0, 1), (1, 2), (2, 3), (3, 4), (4, 5), … ; а, све сагласно са два одновна мендељејевска принципа – принципа минимума промене, и принципа континуитета [Прва фамилија „металних пресека”: „златни” (p=1; q=1), „сребрни” (p=2; q=1), „бронзани” (p=3; q=1), …; Друга фамилија „металних пресека”: „златни” (p=1; q=1), „бакарни” (p=1; q=2), „никлени” (p=1; q=3, …).]
Двоструке вредности бројева које налазимо на дијагонали таблице множења: 0, 4, 12, 24, 40, 60… препознајемо и као вредности једне од катета Диофантових троуглова. Увећане за по 1 те вредности постају хипотенузе истих тих Диофантових троуглова: 1, 5, 13, 25, 41, 61итд. Кад знамо да се друга катета Диофантових троуглова генерише из скупа непарних природих бројева (1, 3, 5, 7, 9, 11…), тада имамо и комплетан запис за све Диофантове троуглове: 0. (1, 0, 1), 1. (5, 4, 3), 2. (13, 12, 5), 3. (25, 24, 7), 4. (41, 40, 9), 5. (61, 60, 11) итд. Надаље, кад знамо све ове везе са златним пресеком, и још знамо да је генетски кôд такође детерминисан златним пресеком (Rakočević, 1998), разумемо и налаз В. Шчербака (V. Shcherbak), према коме кôдирање тзв. четворокодонских аминокиселина „иде” преко Питагорине тројке (5,4,3) (Shcherbak, 1994).
Једна или више трословних речи (кодона) дају значење словима друге азбуке генетског кôда, неком од 20 аминокиселинских молекула. А то давање значења није ништа друго до стварање стереохемијских услова за хемијску реакцију у којој се над свака три молекула нуклеотидних база (које изграђују дугачке нити нуклеинских киселина ДНК и РНК, као на сликама 1 и 2), уграђује по једна нова аминокиселина у протеински ланац.
Молекул дезоксирибонуклеинске киселине (ДНК) је, заправо, макромолекул. Приликом деобе ћелије, његове две спирално увијене нити се расплићу, али тако да свака од њих изграђује нову, себи комплементарну нит. Од једног молекула настају два, па кад се ћелија подели – две новонастале ћелије имају исти генетски запис. Али, не само то. Над једном од две нити молекула ДНК (кодогена нит) синтетизује се једнострука нит информационе рибонуклеинске киселине (iRNK). Она се креће кроз цитоплазму и том приликом се низ њу котрљају ћелијска телашца рибозоми који су, заправо, мале фабрике за производњу протеина.
Механизам је следећи: транспортна рибонуклеинска киселина (tRNK) која допрема аминокиселину, са три своја слова (три нуклеотида) на супротном крају од привезане аминокиселине, тзв. антикодоном, повезује се са три комплементарна нуклеотида iRNK. Тиме су створени стереохемијски услови да се придошла аминокиселина присаједини претходно повезаним аминокиселинама у протеински ланац, како је показано на слици 2. [Као што је општепознато, начин функционисања молекула наслеђа – молекула ДНК – кроз спаривање комплементарних нуклеотида открили су Џејмс Вотсон (James Watson) и Френсис Крик (Francis Crick) 1953. године, за које откриће су 1962. године добили Нобелову награду].
Изненађење у изненађењу
Када сам се први пут, почетком седамдесетих година прошлог века, истраживачки удубио у таблицу генетског кôда, био сам у исто време двапут изненађен и зачуђен. Било је то својеврсно – изненађење у изненађењу. Зачудила ме најпре једноставност те таблице, и чињеница да је један математички израз у потпуности реализован у стварности; али ме још више зачудило то што се људи над овом једноставношћу нису довољно зачудили. Срећом, и то се догодило десетак година касније.
Америчка хемичарка Розмери Свенсон (Rosemarie Swanson), са Универзитета у Тексасу, показала је, 1984. године, да се генетски кôд може свести на греј кôд (Gray code), на одавно познати бинарни машински кôд у којем се наизменично смењују светла и тамна поља. Показала је, наиме, да се 64 генетско-кодне речи (кодони) могу поређати тако да чине један круг у којем се свака следећа реч од претходне разликује тачно за једно слово. С правом је славодобитно закључила да за генетски кôд важе два кључна принципа: принцип минимума промене и принцип континуитета. Права је штета да се том приликом није сетила како су то заправо два изворна мендељејевска принципа, што би јој указало на очекивану могућност да то значи да генетски код „вуче” своје корене управо из периодног система хемијских елемената (ПСЕ).
Наоружан овим сазнањима кренуо сам да тражим задовољење мојој радозналости и зачуђености. Одлучио сам да греј кôд модел и саму таблицу генетског кôда развијем у бинарно-кôдно дрво, 0–63 (слика 3) и преиспитам све битне односе између осам поткласа кодона (осам октета) у оквиру целине као својеврсне класе састављене од 64 кодонске речи. Резултати до којих сам дошао објављени су једним делом, најпре у мојој књизи „Гени, молекул и језик” (Научна књига, 1988), а потом, у дефинитивној форми, у престижном интернационалном часопису (Biosystems, 1998).
Међутим, објављивање књиге је имало своју предисторију. Већ припремљена за штампу остала је да чека неколико месеци током 1987. године, док се нису завршила два моја ненадано добијена, веома повољна студијска боравка у Немачкој, два месеца у Берлину и месец дана у Минхену, у Макс Планк институту, у одељењу за протеинску хемију. Али, оно што је било необично у мом студијском програму, и што је помало изненадило стипендиторе, било је то што сам тражио да се усавршавам истовремено у истраживању генетичких информација и у претраживању научних информација. На тај начин сам се нашао на изворима свих релевантних светских научних база података, и кад сам се вратио кући моја књига је допуњена посебним додатком од читавих педесетак страница, и све су биле посвећене генетском кôду.
Видевши бинарно-кôдно дрво генетског
кôда у мојој књизи, неколико добронамерних,
али и недобронамерних истраживача у
нашој средини, упозорило ме на то да оно
подсећа на исто такво бинарно дрво које
се налази у кинеској „Књизи промена”.
Током боравка у Минхену, у Макс Планк институту, баш тада је ту настајала Међународна база протеинских и нуклеинско-киселинских база – записа свих до тада хемијски анализираних макромолекула ове врсте, тако да сам на лицу места могао да се уверим да мој, хемијски и кодогено заснован, хијејархијски редослед 20 протеинских аминокиселина боље „пасује” од службеног, садржаног у реченим базама података.
Видевши бинарно-кôдно дрво генетског кôда у мојој књизи, неколико добронамерних, али и недобронамерних истраживача у нашој средини, упозорило ме на то да оно подсећа на исто такво бинарно дрво које се налази у једној од најстаријих књига на свету, у кинеској „Књизи промена” (Ји-Ђинг), старој око три хиљаде година. Био сам скептик и, упркос свим упозорењима, у почетку ми није било ни на крај памети да проверим, да погледам о каквој је то староставној књизи реч. Нарочито због тога што сам на предавањимна из филозофије већ одавно био сазнао за изричито негативан Хегелов став према тој књизи.
Изворни кључ живота
До преокрета је дошло кад ми је, једном потпуно случајном околношћу (најпре на немачком, а потом и на енглеском) допала руку књига, за мене тада неочекиваног наслова, „Ји Ђинг и генетски кôд”, с поднасловом „Заборављени кључ живота”. Аутор је био немачки лекар Мартин Шенбергер (Martin Schoenberger), који је одмах након обелодањивања генетског кôда увидео могућу аналогију и везу. [Мартин Шенбергер је књигу објавио 1973. године. Касније сам нашао да је познати молекуларни биолог Гантер Стент (Gunther S. Stent) још 1969. објавио књигу The comming of the golden age у којој је анализирао аналогије и везе између генетског кôда и „Ји Ђинга”, тако да му припада приоритет].
Кинези нису чекали на нас из трећег
миленијума, него су пре три хиљадe
година на основу 64 хексаграма на
шестобитном бинарно-кôдном дрвету
изградили своје писмо и филозофију.
Поново је ђаво дошао по своје! Моје истраживачке обавезе су се удвостручиле. Сада је требало истраживати не само генетски кôд већ и стваралаштво старих народа и класика, а све с циљем да се одгонетне могуће „согласије опште”, како би рекао Његош, и да се види у чему је тајна: да ли је генетски кôд, можда, само један аспект неког универзалног кôда природе и које би биле карактеристике једног таквог кôда?
Силом прилика моја истраживања су морала да крену двема линијама. Једна је била тајна, а друга јавна. Оно што је било на тајној линији ни за живу главу нисам смео да објавим у било којем од званичних научних часописа, јер бих одмах био оптужен за нумерологију. „Али, ако не у часописима, објавићу у књигама, што на српском, што на енглеском језику”, тако сам резоновао. И сада свако може да нађе моју књигу Genetic code as a unique system (Студентски културни центар у Нишу, Бина у Београду, 2007b, стр. 60), ако не у књижарама, а оно на мом сајту (www.rakocevcode.rs), и увери се да златни пресек, савршени и пријатељски бројеви заиста јесу детерминанте генетског кôда – детерминанте секвенце природних бројева 0-63 (табела 3).
[Ако је читаоцу прејак израз детерминанте, може се рећи да аминокиселине кореспондирају с реченим ентитетима–квантитетима, преко логичке релације: „ако–онда”: ако златни пресек линеарне дужи 0–63 пада између бројева 38 и 39, а на тим позицијама се налазе кодони CAA и CAG који кодирају аминокиселину глутамин, онда је она „златна” аминокиселина, на позицији ф^1(фи на први степен); истим поступком се надаље откривају и преостале „златне” аминокиселине, на позицијама од ф^2 до ф^9 (слика 3 у релацији с табелом 3]
Шестобитно бинарно дрво је посебно и специфично, поред свега другог и по томе што је једино на њему збир бројева на двема унутрашњим гранама (два октета) толики да одговара првом пару пријатељских бројева (220 + 284 = 504), а по две суседне гране се допуњују до њиховог збира (504); све заједно, реализује се својеврсни логички квадрат: (0) 220 + 284 = 504; (1) 156 + 348 = 504; (2) 92 + 412 = 504; (3) 28 + 476 = 504. С друге стране, збир бројева првог квартета износи 6, првог октета 28; збир бројева на левој грани (што се подудара са збиром бројева у првој половини таблице ГК), од 0 до 31, износи 496, што је редом реализација прва три савршена броја (6, 28, 496).
Ако, пак, избројимо све бројеве 0-63, па се вратимо натраг (цикличност!), при чему број 0 постаје 127, па све бројеве секвенце 0-127 саберемо, добићемо као резултат 8128, што је заправо четврти савршени број. Након ових увида постаје јасно зашто Кинези нису чекали на нас из трећег миленијума, него су све ово још пре три хиљада година увидели и на основу 64 хексаграма на шестобитном бинарно-кôдном дрвету изградили не само своје писмо и филозофију него податкали и основе своје културе.
Да би се потврдило да све ово има не само кодогени већ и хемијски смисао, требало је „златним” аминокиселинама придружити њихове хемијске комплементе и испитати могуће равнотеже, сагласности и правилности (Rakočević, 1998, Survey 2.1 & 2.2, p. 289). [„Златне” аминокиселине: (F, L, Q, T, P, S, G); „златне” аминокиселине с придруженим хемијаким комплементима: (F-Y, L-A, Q-N, T-M, P-I, S-C, G-V); преостали хемијски некомплементи, шест АК, спакованих у три хемијска пара: (D-E, K-R, H-W.] Ово хемијско спаривање 20 АК у десет парова било је, у науци о генетском кôду, прво предочавање да из хемијске природе 20 протеинских аминокиселина неминовно следе десет аминокиселинских парова (Rakočević, 1998, Biosystems, 46, 283-291).[3]
Број атома у бочним низовима молекула предочених трију класа аминокиселина разликује се тачно за (1 x 6), (2 x 6) и (3 x 6). Наиме, у седам „златних аминокиселина” има тачно 60 атома, а у њихових седам комплемената – исто толико плус (1 x 6), што укупно износи 66; у преосталих шест некомплемената има 66 плус (2 x 6) атома што је укупно 78 атома. Ако „златне аминокиселине” напишемо не редоследом како су скинуте с бинарног дрвета већ редоследом према броју атома које у себи поседују, заједно са њиховим комплементима (G-V, S-C, T-M, P-I, Q-N, L-A, F-Y), и испред њих се додају три пара некомплемената, тада се добија уређен систем протеинских АК, према позицијама њихових атома у ПСЕ, који сам назвао Cyclic Invariant Periodic System, у скраћеној форми CIPS: {H-W, K-R, D-E (G-V [S-C, T-M] P-I), Q-N, L-A, F-Y}.
Како видимо, добијају се четири класе аминокиселина, према хемијској сличности, три класе аланинског стереохемијског типа, и једна класа неаланинског типа (Попов, 1989; Rakočević & Jokić, 1996). У средини се налазе четири халкогене АК (имају кисеоник, или сумор у бочном низу): S-C, T-M; у следећем циклусу налазе се четири АК неаланинског типа: G-V, P-I; следи циклус једине две А АК које поседују карбоксилну групу у бочном низу, заједно са њихова два амидна деривата: D-E, Q-N; потом долазе четири алифатичне аминокиселине: K-R, L-A; коначно, у последњем циклусу налазе се четири ароматичне аминокиселине, хемијски комплексније од алифатичних: H-W, F-Y (упоредити: Rakočević, 2011, Fig. 6, p. 832).
[3] Номенклатура протеинских аминокиселина: аланин (Alanine, A), аргинин (ARginine, R), аспарагин (AsparagiNe, N), аспарагинска киселина (Aspartic acid, D), цистеин (Cysteine, C), глутамин (Glutamine, Q), глутаминска киселина (Glutamic acid, E), глицин (Glycine, G), хистидин (Histidine, H), изолеуцин (Isoleucine, I), леуцин (Leucine, L), лизин (Lysine, K), метионин (Methionine, M), фенилаланин (Phenylalanine, F), пролин (Proline, P), серин (Serine, S), треонин (Threonine, T), триптофан (Tryptophan, W), тирозин (Tyrosine, Y), валин (Valine, V).
Пут којим се теже иде
Незнано како и зашто, тек догодило се то да горе цитирани рад Е. М. Попова (Евгений Митрофанович Попов), из 1989. године, осим мене скоро нико више не цитира, кад је реч о применама на генетски кôд.[4] А, без чињеница саопштених у његовом раду генетски кôд практично није могуће разумети; следствено томе, није могуће разумети постанак и суштину живота у универзуму, па се тиме не може имати ни адекватна биологија, нити медицина.
Па, које су то, онда, чињенице? Ево их, белодано дате на слици 4. На самом почетку дата је формула најједноставније протеинске аминокиселине, глицина: за „главу” те аминокиселине (за аминокиселинску функционалну групу), једну те исту у свих 20 аминокиселина, прикачено је „тело” (бочни низ) од само једног атома водоника. Другим речима, између главе и тела нема ничег уметнутог.
Тек са увидом у постојање четири типа
диверзитета протеинских аминокиселина,
били су створени услови да се уочи веза
између генетског и хемијског кôда, и то
кроз аналогије, које су и више од тога.
Међутим, ако посматрамо следећу аминокиселину (следећу по хемијској сложености), аланин, видимо да је између главе и тела уметнута једна CH2 група. Али, кад се упознамо с радом Е. М. Попова о четири стереохемијска типа протеинских аминокиселина, тада уочавамо да чак још 15 аминокиселина имају исту ту CH2 групу између главе и тела. [Изузетак је само треонин у коме је један атом водиника CH2 групе супституисан CH3 групом (слика 4 друга колона, трећи ред).] То, онда, значи да 16 аминокиселина припада аланинском стереохемијском типу, а само једна (глицин) припада глицинском стереохемијском типу.
Избором глицина, природа је из неугљоводоничности изабрала наједноставнији могући случај; не узима надаље ниједан други, већ прелази на угљоводоничност, и из отвореног угљоводоничног низа узима, такође први могући случај, метил групу (CH3) из наједноставнијег органског молекула, метана, и везује је за главу аминокиселине – настаје аланин. Очекивало би се да следећа за везивање буде етил група (CH3-CH2), али то се не догађа. Природа и овде – из отворености – узима први могући случај, и прелази на полуцикличност[5] (слика 4, трећа колона, први ред).]: „троугао” изопропил групе веже се својим „теменом” за главу аминокиселине, настаје валин; потом се веже својом страницом, па настаје први могући циклични аминокиселински молекул, пролин (слика 4, друга колона, први ред).
[4] Тешко је веровати, али разлог би могао бити у чињеници да тзв. „Запад“ са потцењивањем гледа на руску науку. Тако, озбиљан истраживач ће са неверицом утврдити да се и данас на Западу у исту раван стављају Дмитриј Иванович Мендељејев и Лотар Мајер, кад је реч о периодног систему хемијских елемената. Наводно су „исту ствар“ обојица открили, независно један од другог, иако је само Мендељејев открио Закон периодичности.
[5] Наравно, ни у једном уџбенику органске хемије, на читавом свету, не може се наћи појам „полуцикличност“ молекула, али природа не може да „прескаче“, већ се увек понаша у складу са два природна принципа, које препознајемо као мендељејевске принципе – са принципом минимума промене и принципом континуитета.
Након тога природа прелази на рачвастост. Први случај рачвања отвореног низа могућ је тек са четри атома угљеника: настају две изомерне аминокиселине, леуцин и изолеуцин (слика 4, друга колона, први ред). Са овим су сва четири стереохемијска типа протеинских аминокиселина генерисана: глицински (са глицином), пролински (са пролином), валински (са валином и изолеуцином), аланински, са преосталих 16 аминокиселина. Кључно у стереохемијској геометрији јесте слобода ротације око хемијске везе која повезује главу и тело аминокиселине: у случају глицина стопроцентна слобода ротације; у случају пролина ротација није могућа; у преостала два типа могућа је делимична ротација, код аланинског типа ближа моделу глицина, а код валинског ближа моделу пролина – нијансирање, као и у свему другом, кад је у питању генетски код. Подразумева се да не може бити другачије него да се ова својства аминокиселина одражавају на структуру и функцију протеина, управо од аминокисеина саграђених.
Такође не може бити другачије него то да предочена геометрија јесте само један аспект далеко шире геометрије генетског кода, која се својом укупношћу, а посебно преко златног пресека одражава и на протеине, па тиме и на сâм живот – од најједноставнијих до најсложенијих организама.[6]
*
Кад једном сазнамо о неминовности постојања четири стереохемијска типа протеинских аминокиселина, тада смо на добром (и тешком!) путу да
[6] Примера ради, нобеловац Манфред Ајген (Manfred Eigen), заједно са својим колегом Петером Шустером (Peter Schuster), и сâму Таблицу генетског кода предочио је, деведесетих година прошлог века, у форми тродимензионалне коцке. Розмери Свенсон, у већ цитираном раду (Swanson, 1984) говори о „коцки кодонског пута“. Што се, пак, мене тиче, у својој књизи из 1994. године предочио сам LIGHT (Logical–Information–Geometric–Homomorphic–Topological) модел коцке (Rakočević, 1994) који важи, како за Таблицу, тако и за бинарно дрво генетског кода; модел у коме се интегришу логичка, информациона, геометријска и тополошка суштина стандардног генетског кода. У најновије време (12. јуна 2017.), у часопису „Frontiers in Computational Neuroscience“, објављена је вест о научном резултату [„Scientists discover hidden patterns of brain activity, École polytechnique fédérale de Lausanne (EPFL)“], према коме се информације које се генеришу у можданим неуронима експримирају у форми специфичних геометријских образаца, што је у науци потпуна новост. Моја хипотеза (и предикција!), током вишедеценијског истраживања генетског кода, све време је била та да се укупна геометрија и стереохемија генетског кода морају експримирати у структури и функцији протеина, а то значи и у сâм живот, ма где он био у Универзуму, у чему и јесте смисао трагања за адекватном биологијом и медицином.
увидимо и постојање четири типа диверзитета истих тих – протеинских аминокиселина. Уочавамо да постоје тачно четири аминокиселине у чијем бочном низу се налазе хемијски елементи угљеник и водоник и ниједан други. Кажемо да су то стандардне угљоводоничне аминокиселине. Али такође лако уочавамо да постоје и две нестандарне, такође угљоводоничне:[7] глицин и пролин.[8]
У даљем трагању за постојањем типова диверзитета, уочавамо да постоји тачно осам аминокиселина код којих се функционалне групе из главе аминокиселине пресликавају и у тело, у бочни низ. Преостаје шест аминокиселина код којих нема тог пресликавања. Све заједно, добијамо четири (диверзитетски различите) класе аминокиселина, уређених по хемијским паровима: [(G, P), (A-L, V-I), (C-M, F-Y, H-W), (S-T, D-E, N-Q, K-R)].
Са аспекта разликовања у основном физичко-хемијском својству, две нестандардне угљоводоничне аминокиселине су семиполарне; четри угљоводоничне неполарне; осам аминокиселина код којих постоји речено пресликавање јесу поларне; и, коначно, преосталих шест аминокиселина представљају комбинацију неполарних, поларних и семиполарних. [Семиполарна је трифтофан (W), поларне су хистидин (H) и тирозин (Y), а неполарне су: фениланалин (F), цистеин (C) и метионин (M).]
Тек са увидом у постојање четири типа диверзитета протеинских аминокиселина, били су створени услови да се уочи веза између генетског и хемијског кôда, и то кроз аналогије, које су и више од тога (оне су суштинске карактеристике и једног и другог кôда). Показује се, наиме, да је кодирање у оба кôда засновано на истом моделу. То је систем од две ленеарне једначине чија десна страна код сумирања x + y даје, у првој једначини квадрат броја 5, а у другој квадрат броја 6; у случају разлика x – y резултат је квадрат броја 3 и броја 4.
У стварности то значи следеће: у стандардном генетском кôду постоји тачно 61 аминокиселински кодон (дају значење аминокиселинама) и 3 „стоп” кодона (означавају „стоп” ситуације, тј. прекид у синтези протеина). Од 61 аминокиселинских кодона, 25 се троши на кôдирање 2+4 мање комплексних аминокиселина, оних које у бочном низу садрже само елементе водоник и угљеник (код глицина је само водоник).
С друге стране, 36 кодона се троши на кодирање комплекснијих аминокиселина, њих 6+8, код којих у бочном низу осим угљеника и водоника постоји бар још један елеменат – кисеоник, или азот, или сумпор. Даљи ток дистрибуције је, како следи. Решења линеарне једначине чија је десна страна 25, јесу 8 и 17, а то одговара броју кодона у процесу кодирања: 8 кодона кодира две нестандардне угљоводоничне аминокиселине, а 17 кодона кодира четири стандардне угљоводоничне аминокиселине.
Линеарна једначина чија је десна страна 36 има решења 10 и 26; од тога, 10 кодона се троши на кодирање шест аминокиселина које немају пресликавање глава–тело, а 26 кодона на кодирање оних осам аминокиселина које пресликавање глава–тело поседују.
*
Аналогне ситуације постоје и у хемијском кôду, оваплоћеном у Периодном систему хемијских елемената, кад се ПСЕ посматра до границе стабилности/нестабилности, до полонијума (P-84). У том скупу, 20 моноизотопних елемената одговара стању од 20 некодонских ситуација (20 аминокиселина), 61 вишеизотопни елеменат одговара стању „61 аминокиселински кодон”, а три прекида у стабилној изотопији (технецијум, Tc; прометијум, Pm и полонијум, Po) одговарају трима „стоп“ кодонима у стандардном генетском коду.
Надаље се скуп од 61 вишеизотопног елемента разбија тачно као и у генетском кôду, на 25 и 36. При томе, 25 хемијских елемената су мање стабилни, a 36 су више стабилни. [Мање стабилни, нестабилни, у смислу да сваки од њих поседује бар по један примордијални нестабилни изотоп; више стабилни, стабилни у смислу да ниједан од тих 36 елемената не поседује нестабилне изотопе, већ само стабилне.] Даља дистинкција иде по истим оним двема линеарним једначинама које важе за генестки кôд, и то у следећем смислу: 25 нестабилних елемената, њих осам, налази се у непарним групама периодног система, док се њих 17 налазе у парним групама периодног система, таквог ПСЕ, какав је дат пре 26 година (Rakočević, 1991).
На приближно исти начин изведена је дистинкција у скупу од 36 стабилних елемената, с малим померањем у решењима линеарне једначине; уместо x = 10 и y = 26, овде имамо решења x = 13 и y = 23. У реалности то значи да се 13 стабилних елемената налазе у непарним групама ПСЕ, а 23 у парним. (Детаље видети у чланку Genetic code and periodic system: some analogies на сајту www.rakocevcode.rs)[9]
Овим се завршава доказни поступак у покушају да се одговори и на два потпитања, са почетка овог чланка – зашто баш та четири молекула у изградњи макромолекула ДНК и РНК; и зашто баш тих 20 аминокиселинских молекула у изградњи макромолекула протеина? Одговор је у предоченим јединственостима и једноставностима тих молекула (проистеклих из једноставности атома који их граде), и у кореспонденцији њихових кодирајућих аранжмана са јединственим и једноставним математичким правилностима и законитостима.
Нема те математичке теорије вероватноће којом би се могло показати да све ово, што је горе изложено, јесте случајно. Напротив, све је резултат његошевског „согласија општег”, које се своди на то да су у природи могући само они системи који су детерминисани најбољом могућом – симетријом, пропорцијом и хармонијом, у чијој основи је златни пресек.
[7] Упркос томе што у бочном низу има само водоников атом, глицин је такође угљоводонична аминокиселина, с обзиром на то да у глави поседује угљеник.
[8] Нестандардност пролина потиче отуда што код њега постоје повезаност главе и тела.
[9] Предочено „померање“ за 03 / 13, 23 другачије је од свих других, могућих у датој линеарној једначини, са аспекта важења/неважења принципа сличности и самосличности: (01 / 11, 25), (02 / 12, 24), (03 / 13, 23), (04 / 14, 22), итд.
ИЛУСТРАЦИЈЕ

Преглед 1. Експоненцијални изрази према којима су генерисани природни кодови: лево – хемијски код; и десно – генетски код. Према изразима на левој страни генерише се Таблица периодног система хемијских ееената (ПСЕ); односе се на распоред елемената до границе стабилности, до полонијума, који је 84. елеменат, са 84 протона у језгру аттома. По 9 елемената налазе се у једној групи осам пута; по 4 елемента два пута, и по један елеменат четири пута. Према изразима на десној страни генерише се Таблица генетског кôда (табела 1), са 4 синглета, 16 дублета и 64 триплета; такође и бинарно дрво (слика 3) са 2, 4, 5, 16 и 32 гране респективно; у честом кораку генеришу се 64 кодонска триплета.
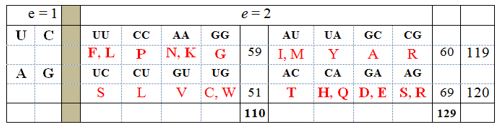
Преглед 2. Генерисање 16 нуклеотидних дублета из четворословне генетско-кодне азбуке, према изразу mn (m = 4; n = 2; e = експонент). Ако је e = 1, тада имао четири синглета, четири базе – две пиримидинске (урацил, U и цитозин, C), са по једим прстеном у молекулу (хемијски једноставније), и две пуринске (аденин, A и глутамин, G), са по два прстена у молекулу (хемијски комплексније). Ако је e = 2, тада имао 16 нуклеотидних дублета. Ако се дублети уреде по хемијској хијерархији (најпре једноставнији UU, CC; потом комплекснији AA, GG), тада видимо да је ову хемијску суштину прати равнотежа броја атома у бочним низовима аминокиселина, које су тим дублетима кодирене (све сагласно Аристотеловој идеји о јединству форме и суштине).
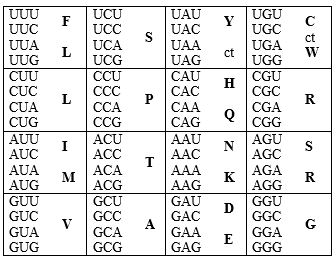
Табела 1. Таблица стандардног генетског кôда. Од три могућа аранжмана (исто слово по 16 пута, респективно – у првој, другој, или трећој позицији) овде је дат онај који најбоље одговара хемијској сличности, како кодона, тако и аминокиселина (Crick, 1968).
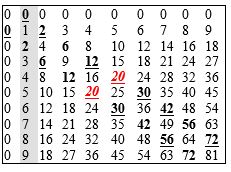
Табела 2. Таблица множења у декадном бројевном систему (са бројем 20 у средини, у форми дублета 20-20).

Табела 3. Секвенца низа природних бројева 0-63. Ако се уреди по октетима, што је случај у ГК (слика 3), тада је та секвенца строго детерминисана првим паром тзв. пријатељских бројева (220 и 284), као и трећим савршеним бројем (496). Осим тога, сума свака два октета допуњује се до суме прва два пријатељска броја, кореспондентно са логичким квадратом: (0) 220 + 284 = 504; (1) 156 + 348 = 504; (2) 92 + 412 = 504; (3) 28 + 476 = 504. Ово овако могуће је само на шесто-битном бинарном дрвету, чиме се потврђује његова уникатност, а тиме и уникатност генетског кода. [У пријатељском пару сума чинилаца првог броја даје други, и обрнуто; а, савршен је онај број који је једнак суми својих чинилаца.]
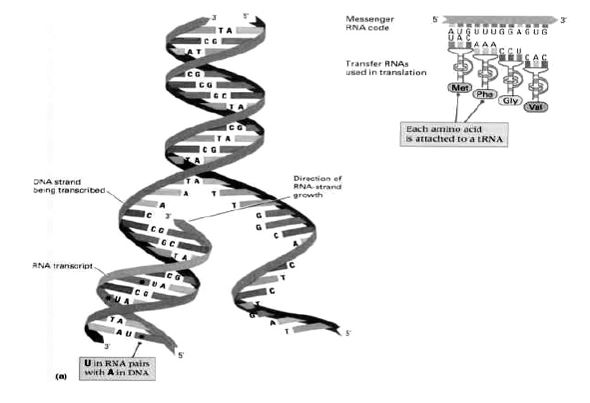
Слика 1. Молекул дезоксирибонуклеинске киселине – макромолекул наслеђа (DNK). Расплитањем две нити при деоби генеративне ћелије, једна нит остаје мајци, а друга, у истоветном запису се предаје ћерки-ћелији; притом свака од две нове нити догради себи недостајућу, комплементарну нит.
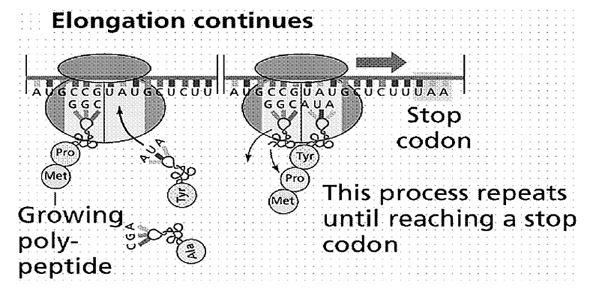
Слика 2. Резултат генетског кодирања: над једном од две нити макромолекула ДНК синтетизује се једнострука нит рибонуклеинске киселине (РНК), низ коју се у цитоплазми ћелије котрљају мала лоптаста телашца – рибозоми. На њима се генерише систем кодон-антикодон (кодони на информационој и антикодони на транспортној РНК), са доношењем одговарајуће аминокиселине (помоћу транспортне РНК). Потом, као на шиваћој машини, пришивају се једна по једна аминокиселина, добија се полипептидни ланац, као основна нит за изградњу макромолекула протеина.
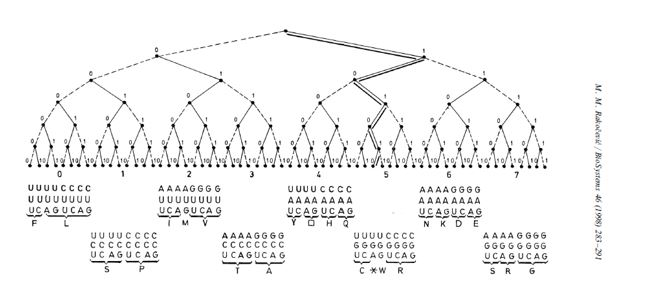
Слика 3. Шестобитно бинарно дрво генетског кода, са распоредом 64 трословне „речи” (кодона), генерисаних из четворословне азбуке (U,C,A,G), од почетне, то јест нулте, UUU (000000), преко хармонијске (у позицији хармонијске средине), са највећом променом, UGA 101010, до последње, на позицији 63 (111111). [Према: M. M. Rakočević, BioSystems 46 (1998), 283–291.]
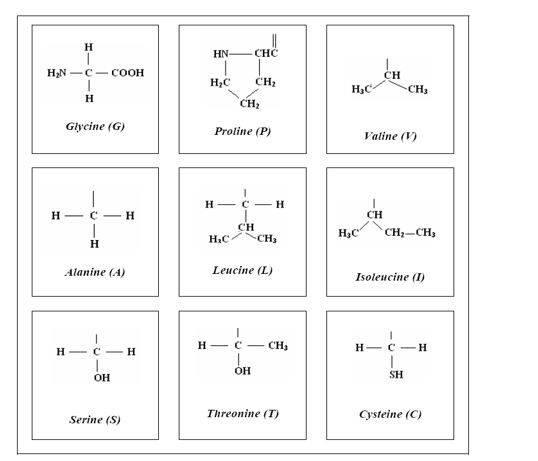
Слика 4. Структурна формула глицина и бочног низа преосталих 19 протеинских аминокиселина. Формуле кореспондирају са четири стереохемијска типа: глицински (са глицином), пролински (са пролином), валински (са валином и изолеуцином), аланински, са преосталих 16 аминокиселина, од којих је овде приказано пет карактеристичних.
РЕФЕРЕНЦЕ
Crick, F.H.C. (1968) The origin of the genetic code, J. Mol. Biol., 38, 367- 379, 1968.
Кедров, Б.М. (1977) Прогнозы Д.И. Менделеева в атомистике, Атомиздат, Москва.
Попов, Е. М. (1989) Структурная организация белков, Наука, Москва.
Rakočević, M.M (1991) The coherence of the chemical and genetic code, Zbornik radova Filozofskog fakulteta u Nišu, Sekcija za hemiju, 2, 1-29 (Tabela 4.1; ova Tabela se nalazi i u knjizi: Rakočević, 1997b, kao Tabela 18).
Ракочевић, М. М. (1988) Гени, молекули, језик, Научна књига, Београд. (www.rakocevcode.rs)
Rakočević, M. M. (1994) Logic of the Genetic Code, Naučna knjiga, Beograd. (www.rakocevcode.rs)
Rakočević, M. M.1(997a). Two classes of the amino acyl-tRNA synthetases in correspondence with the Codon path cube. Bull. Math. Biol. 59, 645-648.
Rakočević, M. M. (1997b) Genetic code as a unique system, Студентски културни центар у Нишу, Бина у Београду. (www.rakocevcode.rs)
Rakočević, M. M. (1998) The genetic code as a Golden mean determined system, Biosystems, 46, 283-291, (www.rakocevcode.rs).
Rakočević, M. M. (2004a) A harmonic structure of the genetic code, Journal of Theoretical Biology, 229, 221-234 (www.rakocevcode.rs)
Rakočević, M. M. (2004b) Further generalization of Golden mean in relation to Euler’s „divine“ equation, FME Transactions (Faculty of Mechanical Engineering, Belgrade, Serbia), 32, 95-98, 2004; arXiv:math/0611095v1[math.GM]. (www.rakocevcode.rs)
Rakočević M. M. (2011) Genetic code as a coherent system, NeuroQuantology, 9 (4), 821-841.
Rakočević, M., Jokić (1996) Four Stereochemical Types of Protein Amino Acids: Synchronic Determination with Chemical Characteristics, Atom and Nucleon Number, J Theor. Biol., 183, 345–349.
Shcherbak, V. I. (1994) Sixty-four Triplets and 20 Canonical Amino Acids of the Genetic Code: the Arithmetical Regularities. Part II. J. Theor. Biol. 166, 475-477.
Spinadel, V. W. de. (1998) From the Golden Mean to Chaos, Buenos Aires.
Spinadel, V.W. de. (1999) The family of metallic means, Visual Mathematics,1, No. 3. (http//members.tripod.com/vismath1/spinadel/).
Swanson, R. (1984) A unifying concept for the amino acid code, Bull. Math. Biol., 46, 187-207.


Zasto postoje 3 stop kodona?
Do odgovora na ovo pitanje doći ćete na sledeći način. Otvorite moj sajt http://www.rakocevcode.rs i u rubrici „pretraga“ otkucajte NQ [Rakočević M. M. (2011) Genetic code as a coherent system, NeuroQuantology, 9 (4), 821-841]. Uzmite u postupak „Table 4“, u relaciji sa „Figure 3“ i „Table 6“. Radi se o tome da aminokislinskih kodona mora biti 25 + 36 (plus 3 stop kodona); 25 kodona kodirajućih za aminokiseline niže kompleksnosti (u bočnom nizu poseduju samo vodonik ili vodonik i ugljenik) i 36 kodona, kodirajućih za aminokiseline više kompleksnosti. Želim Vam pun uspeh.
Miloje M. Rakočević
Dragi gospodine Rakocevic,
dalu ste culi za izjavu Nikole Tesle kojom je opisao mnoge naucnike?
„Mnogi naucnici od samog pocetka ne razmisle dobro i neznaju da od samog pocetka grese!“
Dali ste Vi sigurni u to sta Vi izjavljujete i o cemu pisete i o cemu predajete ?
Sta ako ste pogresili?
To sto ste Vi izjavili je simetralna perfekcija spajanja perfektno simetralnih srodnih hemijskih i fizikalnih i matematickih i bioloskih jedinjenja- spajanje srodnih stanica- inses, po tom principu je napravljena atomska bomba, nasilno spajanje srodnih atoma.
Ja Vam moram nesto reci,
u PRIRODI ne pistoji identican Atom s drugim, Atom je ziv i ima energiju i krece se, u zavisnosti od brzine i velicine kojom se spaja se na polovima s drugim polom drugog atoma koji nije istog magnetskog naboja, princip
Mesec Zemlja Sunce.
Zlatni presek se ne desava tacno u sredini , jer da je po toj vasoj perfekciji nastao zivot, zivot se nebi ni desio.
Za vrsta perfekcije postoji samo u onima koji su dosli na tu ideju da na taj nacin prave atomsko i biolosko oruzje koje ubija i truje, igrate se sa opasnim stvarima i mislite da ste pametni.
Genetska manipulacija je nasilno spajanje srodnih stanica.
Ljudski telo zivotinjsko telo biljno telo je nasilno „smuckano“ jedno s drugim.
Vatikanski naucnici glume pamet i misle da su pametniji od Tvorca i Majke Prirode, a Vi ste ucili po njihovim knjigama i ugledali se na budu, sve laz, to se moze tako ali to nije DOBRO.
Vi kao naucnik imate veliku odgovornost sta cete da izjavljujete i da tvrdite, a ne da slusate na sta Vas nagovara djavo, zasto ovo zasto ono, niste ni svesni koliko ste u pravu sto se tice djavola, on Vam pokaze perfekciju i Vi se zadivite kako je sve jedinstveno perfektno, perfekcija je zlo covece.
Par i par se ne spaja
nepar i nepar se ne spaja
ako hoces da dobijes zdravu materiju
Slepo crevo nije greske prirode
nego genetsko manipulisan crv vatikanskih naucnika kojeg su pustili u prirodu i on se ugnezdio u svim organizmima, zakacio se za creva, parazit koji se hrani od naseg organizma i producira crve kao sto je i on i ubrizgava u probavni sistem iz kojeg dospjevaju i u krv i u kostanu srz jer su osmisljeni i napravljeni i bore se za zivot.
Meso se ne jede, riba se ne jede, kukci se ne jedu, crvi se ne jedu,
Da bi procistio organizam pij caj i jedi samo povrce, sto manje to bolje, treniraj, treniraj misli,
ja sam zdrav ja sam uman
dobro ne ovosi o zlu
zlo se desava samo na Planeti Majci
u coveku jer su se neki uspeotivili Tvorcu.
Ja Vama ne zelim da pametujem jer ovo sto sam ja napisala nista nije moje novootkrice, to su znali mnogi ljudi pre mene, ja sam dosta vremena provodila uz Nikolu Teslu i Svetozara Radisica, bilo bi dobro da se obratite njima a ne Kinezima.
Sretno.