

Одавно је познато да физичко-хемијска својства воде у ћелијама нису иста као у ванћелијском простору, међутим још није нађен убедљив биофизички модел који би објаснио ову разлику. Мистериозно понашање воде у биолошким системима произвело је низ „мистичних” теорија.

Проф. др Павле Р. Анђус
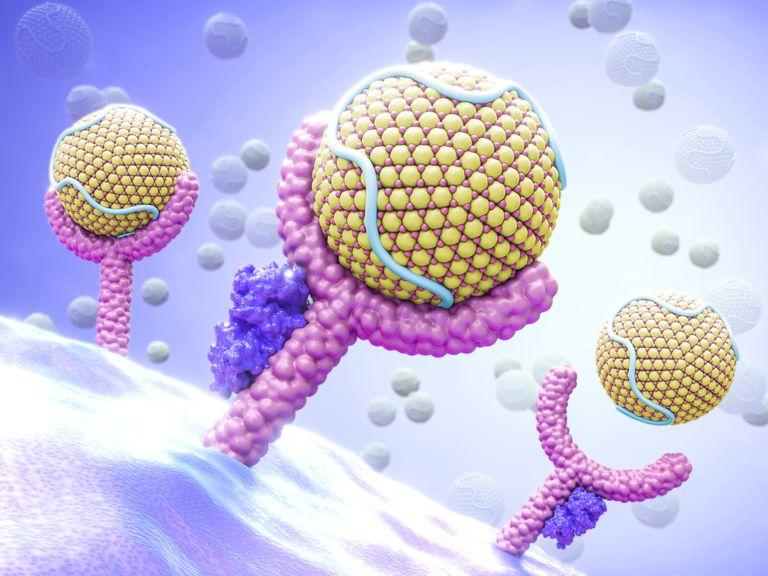
Нобелова нагарада 2003. године за хемију подељена је између Питер Агрија (Peter Agre) са Џон Хопкинс Универзитета из Балтимора и Род Маккинона (Rod MacKinnon) са Рокфелеровог универзитета у Њујорку, а за истраживања структуре и функције протеинских канала за транспорт воде и јона кроз ћелијску мембрану. Док су канали за јоне познати још од 60-тих година и већ су били предмет Нобелове награде (в. доле) канали за воду откривени су тек у последњој деценији прошлог века. За то је заслужан др Агри који их је назвао аквапоринима и објаснио како се селективност за воду у односу на јоне постиже системом водоничних веза у самој пори канала.
Вест о овој Нобеловој награди дочекао сам на почетку своје академске каријере са усхићењем и осећајем носталгије. Одмах сам се сетио семинара 1992. године у угледној Маринско-биолошкој лабораторији Вудс Хол у САД, када сам сматран чудаком, јер правим проблем од нечег тако баналног и добро познатог као што је ћелијски транспорт воде. Откуда таква реакција?
Биофизика мембранског транспорта била је дуже од три деценије под доминацијом истраживања јонског транспорта. Развој ове области инициран је педесетих година прошлог века истраживањима јонске основе нервног импулса нобеловаца Алана Хочкина (Alan Hodgkin) и Ендрју Хакслија (Andrew Huxley), а посебан развој уследио је открићем нове технике наметнуте волтаже на фрагменту мембране (patch-clamp) чији су изумитељи такође нобеловци, немачки научници Берт Сакман и Ервин Нехер. Ова техника је омогућила електрофизиолошку експлорацију појединачних макромолекулских комплекса протеина тј. јонских вратница и канала уграђених у мембране како биљних тако и животињских ћелија. Овај нови експериментални приступ довео је до значајног развоја физиологије ексцитабилних ћелија и неурофизиологије уопште.
Све до краја осамдесетих година сматрало
се да се молекули воде транспортују заједно
са јонима кроз трансмембранске канале.
Међутим, у овом периоду транспорт молекула воде, иако од суштинске важности за електро-осмотску равнотежу ћелија, није привлачио посебну пажњу истраживача. Све до краја осамдесетих година сматрало се да се молекули воде транспортују заједно са јонима кроз трансмембранске канале. Предложено је и више модела пасивног проласка молекула воде кроз слободне просторе у мембранском липидном двослоју. Да ли због „шасивнох” односа науке према пасивном транспорту воде или због фасцинације јонским каналима, но тек почетком деведесетих година, открићем CHIP (channel forming membrane integral protein) макромолекула у еритроцитима, разоткривена је, како у животиња тако и у биљака цела популација мембранских канала – аквапорина, селективних за тако животно важан, наизглед једноставан, али још увек недовољно испитан молекул какав је вода.
Мембрана (Википедија)
Посебан биофизички проблем представља сама структура тј. уређеност воде у живим системима. На овом пољу поломљена су многа научна копља. Одавно је познато да физичко-хемијска својства воде у ћелијама нису иста као у ванћелијском простору, међутим још није нађен убедљив биофизички модел који би објаснио ову разлику. Мистериозно понашање воде у биолошким системима произвело је низ „мистичних” теорија, почев од оне Бориса Дерјагина (одбачене тек након десетак година) о полимеризацији воде (polywate) која је у своје време уздрмала многе озбиљне научне ауторитете, па до наизглед неозбиљне, али патентиране теорије о кластерима воде за које се сматра да су присутни само у амнионској течности фетуса или у лековитој води из светилишта у Лурду! Ипак, решење загонетке понашања „биолошке” воде морало је бити на самој ћелијској мембрани.
У Институт за општу и физичку хемију стигао сам 1981. године, као тек дипломирани студент молекуларне биологије и физиологије. Институтом је руководио професор Душан Вучелић коме је погодовало да има биолога у својој мултидисциплинарној екипи биофизичара, и ја сам се задесио у „право време на правом месту”. Проблематика којом је требало да почнем да се бавим била је управо биофизика мембранског транспорта воде.
У НМР лабораторији Института, применом оригиналне методе др Слободана Мацуре (тада већ на постдокторским студијама у иностранству, а сада директора Одељења за магнетну резонанцу Мејо клинике у Рочестеру, САД), мерили смо транспорт воде кроз ћелијску мембрану слатководне алге из породице харофита, Characeae. Дошли смо до занимљивих резултата везаних за температурску зависност трансмембранског транспота воде у ове алге, који је показивао неочекивано нелинеарно понашање. Наиме, брзина трансмембранске измене воде нагло је опадала у околини 12 и 300 C.

Слободан Мацура (Википедија)
Некако паралелно са овим налазима дошао сам, претрагом литературе, до радова проф. Валтер Дрост-Хансена са Универзитета у Мајамију о фазним прелазима воде на површинама. Радило се о биофизичком моделу који описује реорганизацију молекула воде на површинама и промене структуре водених слојева управо на карактеристичним температурама максимума полувремена трансмембранске измене воде у нашим експериментима. Штавише, наши налази са транспортом воде у Characeae показивали су изразиту сличност са термалним профилима добијенима за фазне прелазе воде у вештачким физичко-хемијским системима.
Десило се да је сарадник Дрост-Хансена, Џејмс Kлег (James Clegg) сазнао за наш рад и тако је отпочела писмена комуникација и размена искустава и резултата која се 1990. године завршила позивом да одржим предавање на конгресу Америчког хемијског друштва у Сан Дијегу. Нажалост, новца за пут није било и сам Дрост-Хансен се понудио да презентира у моје име наше резултате, и тако и би!
Управо је мембранска вода могла бити
један од важних структурних фактора
у вези са општом функцијом мембране.
У каснијим електрофизиолошким експериментима открили смо да и крива температурске зависности мембранске разлике јонских потенцијала алге Chara показује изразите преломе на температурама 15 и 300 C, баш као и водени фазни прелази. Врло слична температурска зависност указивала је на општи механизам у основи термалних промена мембранске грађе и функције, као што су мембрански потенцијал одн. трансмембрански транспорт воде. Управо је мембранска вода могла бити један од важних структурних фактора у вези са општом функцијом мембране.
Функционални значај мембранске воде драматично је илустрован у електрофизиолошким експериментима замене обичне воде тешком водом (деутеријум оксид, D2O). Ефекат додатка само једног неутрона у молекул воде (замена водоника деутеријумом) био је изненађујући – D2O је изазивала наглу мембранску деполаризацију налик ексцитаторним процесима биљне ћелије. Ово откриће је отворило цело ново поље истраживања улоге воде у структури и функцији јонских канала, при чему изомер D2O има улогу водене структурно-функционалне сонде.
На пољу ових истраживања нарочито плодну сарадњу имали смо са екипом проф. Генрика Н. Берестовског из Института ћелијске биофизике Научног центра у Пушћину под Москвом. Сарадници ове лабораторије чак су нам и помогли да формирамо у Београду нашу електрофизиолошку лабораторију са patch-clamp инструментацијом (в. горе) прву те врсте у земљи.
Тада смо први пут претпоставили
постојање посебних мембранских канала
који би проводили само воду насупрот
каналима за катјоне проводним и за воду.
Међутим, данас након доделе Нобелове награде за откриће аквапорина с носталгијом се присећам једне посебне серије експеримената која је чак била предмет мог магистарског рада. Употребом различитих фармаколошких блокатора транспорта кроз ћелијску мембрану на карактеристичним температурама тада смо уочили да постоје диференцијални ефекти на транспорт воде који су могли бити објашњени постојањем два различита мембранска канала за воду. Тада смо, у ствари, први пут претпоставили постојање посебних мембранских канала који би проводили само воду насупрот каналима за катјоне проводним и за воду. Посебни канали за воду били би „уско грло” транспорта воде у околини 15 и 300 C, што је могло да објасни нагли пад транспорта воде на тим температурама.

Станко Стојилковић (Википедија)
Предложено је да је ово успорење протока воде кроз мембрану управо условљено фазним прелазима мембранске воде по Дрост-Хансену. Другој врсти термалних промена, у структури мембранских липида, предложена је улога у увођењу ћелијске мембране у оптимално стање физичко-хемијског медијума за организацију протеинских трансмембранских канала. Показало се да транспорт воде и проводљивост јонских канала за калијум реагују попут система са термосензорима за границе оптималног биокинетичког опсега.
„Добро је док дају Нобелову награду за
нашу област истраживања, а правде у
овој струци ионако никад није ни било…”
(Станко Стојилковић)
Улогу молекуларно-биолошког термосензора могла је одиграти мембранска (површинска) вода са својим фазним прелазима у уским областима на границама биокинетичког опсега температура. Ову хипотезу и резултате који су јој претходили публиковали смо 1991. године у домаћем часопису Periodicum biologorum. Следећа година била је преломна у мојој каријери –добио сам стипендију да похађам престижни курс неуробиологије у Вудс Холу, САД, где сам одржао „уврнути” семинар с почетка овог текста. Међутим, нешто касније те године Питер Агри у часопису Science публикује први рад о аквапоринима. Остало је историја…
Kада сам пријатељу и колеги Станку Стојилковићу, из NIH-а у Вашингтону испричао ову исту причу добио сам одговор: „Добро је док дају Нобелову награду за нашу област истраживања, а правде у овој струци ионако никад није ни било…” Ипак, моје срце је на месту, јер знам да сам тих давних година дотакао једну од безбројних, али не и лако доступних истина природе.





